Schistosoma mansoni és una espècie de platihelmint trematode. Són paràsits dels humans, causant la malaltia coneguda com a esquistosomiasi o bilharzia.
Es distingeixen d'altres trematodes per tenir els sexes separats (la majoria dels trematodes són hermafrodites). El cicle de vida inclou a dos hostes: l'home (hoste definitiu) i un mol·lusc (hoste intermediari).[1][2] És l'únic agent etiològic causant de bilharzia a Amèrica Llatina.
De la mateixa manera que la resta dels trematodes, Schistosoma mansoni té un cos aplanat, no segmentat, revestit per una estructura sincicial anomenada tegument, el qual és metabòlicament actiu en l'absorció de nutrients. Tenen un aparell digestiu incomplet, sense anus.
Schistosoma mansoni és una espècie de platihelmint trematode. Són paràsits dels humans, causant la malaltia coneguda com a esquistosomiasi o bilharzia.
Es distingeixen d'altres trematodes per tenir els sexes separats (la majoria dels trematodes són hermafrodites). El cicle de vida inclou a dos hostes: l'home (hoste definitiu) i un mol·lusc (hoste intermediari). És l'únic agent etiològic causant de bilharzia a Amèrica Llatina.
Krevnička střevní (Schistosoma mansoni) je parazitický helmint z čeledi Schistosomatidae, který způsobuje tzv. střevní schistosomózu. Při onemocnění jsou většinou zasažena játra a tlusté střevo. Mezi definitivní hostitele patří hlavně člověk, ale také další primáti a hlodavci. Mezihostitelem jsou plži z rodu Biomphalaria.[1] Krevnička střevní se vyskytuje v Africe (včetně Madagaskaru a Středního Východu) a v Jižní Americe. Byla pojmenována podle skotského lékaře sira Patricka Mansona.
U krevniček střevních stejně jako u ostatních schistosom nacházíme výrazný pohlavní dimorfismus. Samci mají rozměry 6–12×1 mm, samice 10–16×0, 2 mm. Samice jsou tmavší (jejich trávicí soustava obsahuje pigment hemozoin). Samice je umístěna v břišní rýze samce (tzv. canalis gynaecophorus). Krevničky střevní mají slepou trávicí soustavu, jejich jícen se větví na dvě části, které se spojují v jedno střevo. Jejich vajíčka mají laterálně umístěný trn.[1]
Krevničky střevní mají 8 párů chromozómů. Samice jsou pohlavím heterogametickým (ZW), samci pak homogametickým (ZZ). Jejich genom je asi 270 MB velký, byl poprvé sekvenován v roce 2000. Nyní je popsáno nejméně 11 809 genů. U S. mansoni nacházíme expanzi genů pro proteázy a pro lipidový anabolismus, což odpovídá jejich parazitickému způsobu života.
Životní cyklus je typický pro všechny schistosomy (potažmo i motolice). Z vajíčka se vylíhne obrvené miracidium, které penetruje do vhodného vodního plže (u krevničky střevní se jedná o plže rodu Biomphalaria – výjimkou je případ z Venezuely, kde byla S. mansoni objevena i u suchozemského plže Achatina fulica). V plži dochází k asexuálnímu množení. Vyvíjí se sporocysty a cerkárie, které opouštějí mezihostitele. Furkocerkárie (tj. cerkárie s vidličnatým ocáskem) penetrují do kůže definitivního hostitele. Signálem pro penetraci jsou mastné kyseliny na povrchu kůže. Vniknutí do kůže je umožněno penetračními žlázami. Během tohoto procesu cerkárie odvrhuje ocásek a přeměňuje se v schistosomulu.
Několik dnů zůstávají v kůži, poté se dostává do oběhového systému, ve kterém putují přes plíce a srdce až do cév vrátnicového oběhu a střevního mesenteria, kde čekají na opačné pohlaví. Po spárování začne samička produkovat vajíčka (přibližně 300 za den). Právě vajíčka s trnem jsou hlavním patogenním agens, která při cestě z hostitelova těla porušují tkáně a poškozují orgány.[1] Krevničky střevní bojují proti imunitnímu systému hostitele několika způsoby:
1. Mají povrchový glykokalyx, který svlékají, a tím se stávají méně rozpoznatelné pro imunitní systém
2. Mají tegument, velmi odolný povrch těla
3. Produkují antioxidační proteiny (např. superoxid dismutázy), které je chrání před účinkem superoxidu produkovaného imunitním systémem
Krevničkou střevní je celosvětově nakaženo asi 83,31 milionů lidí. Je přítomna nejvíce na území Afriky a Střední a Jižní Ameriky.
První příznaky schistosomózy (dříve také nazývané bilharzióza podle Theodora Bilharze) se vyskytují 4 až 6 týdnů po nákaze, avšak v mnoha případech se symptomy vůbec neobjeví. Nejdříve se objevuje vyrážka v místě penetrace krevničky, tzv. cerkáriová dermatitis, která intenzivně svědí. Dochází i ke vzniku edémů a ke zduření lymfatických žláz. Po 14 dnech tyto příznaky většinou vymizí.
Akutní formou (2–16 týdnů po infekci) je horečka Katayama, která je doprovázená bolestmi končetin, hlavy, břicha, průjmy (někdy krvavými), únavností a zduřením jater, sleziny a lymfatických žláz.
V chronickém stádiu (3–6 měsíců po nákaze) dochází k zánětlivým reakcím kolem vajíček v různých orgánech, nejvíce v játrech a ve střevě, ale i ve slezině, plicích, srdci, ledvinách nebo centrální nervové soustavě. Tvoří se periportální fibróza. Může se vyskytnout nadměrné zvětšení orgánů (př. hepatosplenomegalie, tj. zvětšení jater a sleziny). Jako komplikace se může objevit i nádorové onemocnění. Závažnost infekce je ovlivňována délkou trvání, počtem usídlených krevniček a jejich lokalizací a celkovým imunitním stavem hostitele.
Krevnička střevní (stejně jako krevnička močová a krevnička jaterní) může v některých případech způsobovat schistosomózu pohlavního traktu žen (Female genital schistosomiasis, FGS), při které dochází k infekci vejcovodů nebo vaječníků vedoucí ke sterilitě. Infekce může zasáhnout jak vnější, tak vnitřní reprodukční orgány.[2]
Diagnóza infekce je založená na najití vajíček S. mansoni ve stolici. Používají se také další metody založené na rozeznávání protilátek (ELISA, APIA = alkaline phosphatase immunoassay; COP = CircumOval Precipitation test). K léčbě schistosomózy se používají anthelmintika ze skupiny chinolových derivátů – praziquantel a oxaminiquin.
(Použita anglická Wikipedie)
Krevnička střevní (Schistosoma mansoni) je parazitický helmint z čeledi Schistosomatidae, který způsobuje tzv. střevní schistosomózu. Při onemocnění jsou většinou zasažena játra a tlusté střevo. Mezi definitivní hostitele patří hlavně člověk, ale také další primáti a hlodavci. Mezihostitelem jsou plži z rodu Biomphalaria. Krevnička střevní se vyskytuje v Africe (včetně Madagaskaru a Středního Východu) a v Jižní Americe. Byla pojmenována podle skotského lékaře sira Patricka Mansona.
Schistosoma mansoni is a water-borne parasite of humans, and belongs to the group of blood flukes (Schistosoma). The adult lives in the blood vessels (mesenteric veins) near the human intestine. It causes intestinal schistosomiasis (similar to S. japonicum, S. mekongi, S. guineensis, and S. intercalatum). Clinical symptoms are caused by the eggs. As the leading cause of schistosomiasis in the world, it is the most prevalent parasite in humans. It is classified as a neglected tropical disease. As of 2021, the World Health Organization reports that 236.6 million people have schistosomiasis and most of it is due to S. mansoni. It is found in Africa, the Middle East, the Caribbean, Brazil, Venezuela and Suriname.[1]
Unlike other flukes (trematodes) in which sexes are not separate (monoecious), schistosomes are unique in that adults are divided into males and females, thus, gonochoric. However, a permanent male-female pair, a condition called in copula, is required to become adults; for this, they are considered as hermaphrodites.
The life cycle of schistosomes includes two hosts: humans as definitive hosts, where the parasite undergoes sexual reproduction, and snails as intermediate hosts, where a series of asexual reproduction takes place. S. mansoni is transmitted through water, where freshwater snails of the genus Biomphalaria act as intermediate hosts. The larvae are able to live in water and infect the hosts by directly penetrating the skin. Prevention of infection is done by improved sanitation and killing the snails. Infection is treated with praziquantel.
S. mansoni was first noted by Theodor Maximillian Bilharz in Egypt in 1851, while discovering S. haematobium. Sir Patrick Manson identified it as unique species in 1902. Louis Westenra Sambon gave the name Schistosomum mansoni in 1907 in honour of Manson.[2][3][4]
Schistosomes, unlike other trematodes, are long and cylindrical worms and are sexually dimorphic. The male S. mansoni is approximately 1 cm long (0.6–1.1 cm)[5] and is 0.1 cm wide. It is white, and it has a funnel-shaped oral sucker at its anterior end followed by a second pediculated ventral sucker. The external part of the worm is composed of a double bilayer, which is continuously renewed as the outer layer, known as the membranocalyx, and is shed continuously.[6] The tegument bears a large number of small tubercules. The suckers have small thorns in their inner part as well as in the buttons around them. The male genital apparatus is composed of 6 to 9 testicular masses, situated dorsally. There is one deferent canal beginning at each testicle, which is connected to a single deferent that dilates into a reservatory, the seminal vesicle, located at the beginning of the gynaecophoric canal. The copula happens through the coaptation of the male and female genital orifices.[7]
The female has a cylindrical body, longer and thinner than the male's (1.2 to 1.6 cm long by 0.016 cm wide). It has the general appearance of a roundworm. The female parasite is darker, and it looks gray. The darker color is due to the presence of a pigment (hemozoin) in its digestive tube.[8] This pigment is derived from the digestion of blood. The ovary is elongated and slightly lobulated and is located on the anterior half of the body. A short oviduct conducts to the ootype, which continues with the uterine tube. In this tube it is possible to find 1 to 2 eggs (rarely 3 to 4) but only 1 egg is observed in the ootype at any one time. The genital pore opens ventrally. The posterior two-thirds of the body contain the viteline glands and their winding canal, which unites with the oviduct a little before it reaches the ootype.[9]
The digestive tube begins at the anterior extremity of the worm, at the bottom of the oral sucker. The digestive tube is composed of an esophagus, which divides in two branches (right and left) and that reunite in a single cecum. The intestines end blindly, meaning that there is no anus.
S. mansoni and other schistosomes are the only flukes or flatworms that exhibit sex separation as they exist as male and female individuals as in dioecious animals.[10][11] However, they are not truly dioecious since the adults live in permanent male-female pairs, a condition called in copula. Although they can be physically separated, [12] isolated females cannot grow into sexually-mature adults.[13][14] In copula starts in the liver only after which they can move to their final habitation, the inferior mesenteric veins. Individual females cannot enter the mesenteric veins.[15] Sex organs, the gonads, are also incompletely separated and are interdependent between sexes. Egg-making organ, the vitelline gland in female does not develop in the absence of male.[16] Male gametes, spermatozoa, are present in the oviduct.[17] In males, there are rudimentary ovaries, oviduct, and oocytes (developing female gametes),[18] as well as vitelline cells.[19] Males also possess the genes for hermaphroditism in flukes.[20] Thus, they are technically hermaphrodites.[18][20]
The eggs are oval-shaped, measuring 115–175 µm long and 45–47 µm wide, and ~150 µm diameter on average. They have pointed spines towards the broader base on one side, i.e. lateral spines. This is an important diagnostic tool because co-infection with S. haematobium (having a terminal-spined eggs) is common, and they are hard to distinguish.[21] When the eggs are released into the water, a lot of them are immature and unfertilised so that they do not hatch. When the eggs are larger than 160 µm in diameter, they also fail to hatch.[22][23]
The miracidium (from the Greek word μειράκιον, meirakion, meaning youth) is pear-shaped, and gradually elongates as it ages. It measures about 136 μm long and 55 μm wide. The body is covered by anucleate epidermal plates separated by epidermal ridges. The epidermal cells give off numerous hair-like cilia on the body surface. There are 17–22 epidermal cells. Epidermal plate is absent only at the extreme anterior called apical papilla, or terebratorium, which contains numerous sensory organelles.[24] Its internal body is almost fully filled with glycogen particles and vesicles.[25]
The cercaria has a characteristic bifurcated tail, classically called furcae (Latin for fork); hence, the name (derived from a Greek word κέρκος, kerkos, meaning tail). The tail is highly flexible and its beating propels the cercaria in water.[26] It is about 0.2 mm long and 47 μm wide, somewhat loosely attached to the main body. The body is pear-shaped and measures 0.24 mm in length and 0.1 mm in width.[27] Its tegument is fully covered with spine. A conspicuous oral sucker is at the apex. As a non-feeding larva, there are no elaborate digestive organs, only oesophagus is distinct. There are three pairs of mucin glands connected to laterally to the oral sucker at the region of the ventral sucker.[28][29]
Developing Schistosoma mansoni worms that have infected their definitive hosts, prior to the sexual pairing of males and females, require a nutrient source in order to properly develop from cercariae to adults. The developing parasites lyse host red blood cells to gain access to nutrients and also makes its own fungi from its waste it is hard to detect; the hemoglobin and amino acids the blood cells contain can be used by the worm to form proteins.[30] While hemoglobin is digested intracellularly, initiated by salivary gland enzymes, iron waste products cannot be used by the worms, and are typically discarded via regurgitation.[31]
Kasschau et al. (1995) tested the effect of temperature and pH on the ability of developing S. mansoni to lyse red blood cells.[30] The researchers found that the parasites were best able to destroy red blood cells for their nutrients at a pH of 5.1 and a temperature of 37 °C.[30]
Schistosoma mansoni is locomotive in primarily two stages of its life cycle: as cercariae swimming freely through a body of freshwater to locate the epidermis of their human hosts, and as developing and fully-fledged adults, migrating throughout their primary host upon infection.[31] Cercariae are attracted to the presence of fatty acids on the skin of their definitive host, and the parasite responds to changes in light and temperature in their freshwater medium to navigate towards the skin.[32] Ressurreicao et al. (2015) tested the roles of various protein kinases in the ability of the parasite to navigate its medium and locate a penetrable host surface.[32] Extracellular signal-regulated kinase and protein kinase C both respond to changes in medium temperature and light levels, and the stimulation of p38 mitogen-activated protein kinase, associated with recognition of parasite host surface, results in a glandular secretion that deteriorates the host epidermis, and allows the parasite to burrow into its host.
The parasite's nervous system contains bilobed ganglia and several nerve cords which splay out to every surface of the body; serotonin is a transmitter distributed widely throughout the nervous system and plays an important role in nervous reception, and stimulating mobility.[33]
After the eggs of the human-dwelling parasite are emitted in the faeces and into the water, the ripe miracidium hatches out of the egg. The hatching happens in response to temperature, light and dilution of faeces with water. The miracidium searches for a suitable freshwater snail belonging to the genus Biomphalaria. In South America, the principal intermediate host is Biomphalaria glabrata, while B. straminea and B. tenagophila are less common.[34] A land snail Achatina fulica was reported in 2010 to act as a host in Venezuela.[35] In Africa, B. glabratra, B. pfeifferi, B. choanomphala and B. sudanica act as the hosts;[36] but in Egypt, the main snail host is B. alexandrina.[37]
Miracidia directly penetrate the soft tissue of snail. Inside the snail, they lose their cilia and develop into mother sporocysts. The sporocysts rapidly multiply by asexual reproduction, each forming numerous daughter sporocysts. The daughter sporocysts move to the liver and gonads of the snail, where they undergo further growth.[38] Within 2–4 weeks, they undergo metamorphosis and give rise to fork-tailed cercariae. Stimulated by light, hundreds of cercariae penetrate out of the snail into water.[39]
The cercaria emerge from the snail during daylight and they propel themselves in water with the aid of their bifurcated tail, actively seeking out their final host. In water, they can live for up to 12 hours, and their maximum infectivity is between 1 and 9 hours after emergence.[40] When they recognise human skin, they penetrate it within a very short time. This occurs in three stages, an initial attachment to the skin, followed by the creeping over the skin searching for a suitable penetration site, often a hair follicle, and finally penetration of the skin into the epidermis using cytolytic secretions from the cercarial post-acetabular, then pre-acetabular glands. On penetration, the head of the cercaria transforms into an endoparasitic larva, the schistosomule. Each schistosomule spends a few days in the skin and then enters the circulation starting at the dermal lymphatics and venules. Here, they feed on blood, regurgitating the haem as hemozoin.[41] The schistosomule migrates to the lungs (5–7 days post-penetration) and then moves via circulation through the left side of the heart to the hepatoportal circulation (>15 days) where, if it meets a partner of the opposite sex, it develops into a sexually mature adult and the pair migrate to the mesenteric veins.[42] Such pairings are monogamous.[43]
Male schistosomes undergo normal maturation and morphological development in the presence or absence of a female, although behavioural, physiological and antigenic differences between males from single-sex, as opposed to bisex, infections have been reported. On the other hand, female schistosomes do not mature without a male. Female schistosomes from single-sex infections are underdeveloped and exhibit an immature reproductive system. Although the maturation of the female worm seems to be dependent on the presence of the mature male, the stimuli for female growth and for reproductive development seem to be independent from each other.
The adult female worm resides within the adult male worm's gynaecophoric canal, which is a modification of the ventral surface of the male, forming a groove. The paired worms move against the flow of blood to their final niche in the mesenteric circulation, where they begin egg production (>32 days). The S. mansoni parasites are found predominantly in the small inferior mesenteric blood vessels surrounding the large intestine and caecal region of the host. Each female lays approximately 300 eggs a day (one egg every 4.8 minutes), which are deposited on the endothelial lining of the venous capillary walls.[44] Most of the body mass of female schistosomes is devoted to the reproductive system. The female converts the equivalent of almost her own body dry weight into eggs each day. The eggs move into the lumen of the host's intestines and are released into the environment with the faeces.
Schistosoma mansoni has 8 pairs of chromosomes (2n = 16)—7 autosomal pairs and 1 sex pair. The female schistosome is heterogametic, or ZW, and the male is homogametic, or ZZ. Sex is determined in the zygote by a chromosomal mechanism. The genome is approximately 270 MB with a GC content of 34%, 4–8% highly repetitive sequence, 32–36% middle repetitive sequence and 60% single copy sequence. Numerous highly or moderately repetitive elements are identified, with at least 30% repetitive DNA. Chromosomes range in size from 18 to 73 MB and can be distinguished by size, shape, and C banding.[45]
In 2000, the first BAC library of Schistosome was constructed.[46] In June 2003, a ~5x whole genome shotgun sequencing project was initiated at the Sanger Institute.[47] Also in 2003, 163,000 ESTs (expressed sequence tags) were generated (by a consortium headed by the University of São Paulo) from six selected developmental stages of this parasite, resulting in 31,000 assembled sequences and an estimated 92% of the 14,000-gene complement.[48]
In 2009 the genomes of both S. mansoni and S. japonicum were published, with each describing 11,809 and 13,469 genes, respectively. S. mansoni genome has increased protease families and deficiencies in lipid anabolism; which are attributed to its parasitic adaptation. Proteases included the invadolysin (host penetration) and cathepsin (blood-feeding) gene families.[49][50]
In 2012, an improved version of the S. mansoni genome was published, which consisted of only 885 scaffolds and more than 81% of the bases organised into chromosomes.[51]
In 2019, Ittiprasert, Brindley and colleagues employed programmed CRISPR/Cas9 knockout of the gene encoding the T2 ribonuclease of the egg of Schistosoma mansoni, advancing functional genomics and reverse genetics in the study of schistosomes, and platyhelminths generally <https://doi.org/10.7554/eLife.41337>.
Schistosome eggs, which may become lodged within the hosts tissues, are the major cause of pathology in schistosomiasis. Some of the deposited eggs reach the outside environment by passing through the wall of the intestine; the rest are swept into the circulation and are filtered out in the periportal tracts of the liver, resulting in periportal fibrosis. Onset of egg laying in humans is sometimes associated with an onset of fever (Katayama fever). This "acute schistosomiasis" is not, however, as important as the chronic forms of the disease. For S. mansoni and S. japonicum, these are "intestinal" and "hepatic schistosomiasis", associated with formation of granulomas around trapped eggs lodged in the intestinal wall or in the liver, respectively. The hepatic form of the disease is the most important, granulomas here giving rise to fibrosis of the liver and hepatosplenomegaly in severe cases. Symptoms and signs depend on the number and location of eggs trapped in the tissues. Initially, the inflammatory reaction is readily reversible. In the latter stages of the disease, the pathology is associated with collagen deposition and fibrosis, resulting in organ damage that may be only partially reversible.[52]
Granuloma formation is initiated by antigens secreted by the miracidium through microscopic pores within the rigid egg shell, and the immune response to granuloma, rather than the direct action of egg antigens, causes the symptoms.[53] The granulomas formed around the eggs impair blood flow in the liver and, as a consequence, induce portal hypertension. With time, collateral circulation is formed and the eggs disseminate into the lungs, where they cause more granulomas, pulmonary arteritis and, later, cor pulmonale. A contributory factor to portal hypertension is Symmers' fibrosis, which develops around branches of the portal veins. This fibrosis occurs only many years after the infection and is presumed to be caused in part by soluble egg antigens and various immune cells that react to them.[54]
Recent research has shown that granuloma size is consistent with levels of IL-13, which plays a prominent role in granuloma formation and granuloma size. IL-13 receptor α 2 (IL-13Rα2) binds IL-13 with high affinity and blocks the effects of IL-13. Thus, this receptor is essential in preventing the progression of schistosomiasis from the acute to the chronic (and deadly) stage of disease. Synthetic IL-13Rα2 given to mice has resulted in significant decreases in granuloma size, implicating IL-13Rα2 as an important target in schistosomiasis.[55]
S. mansoni infection often occurs alongside those of viral hepatitis, either hepatitis B virus (HBV) or hepatitis C virus (HCV). This is due to high prevalence of schistosomiasis in areas where chronic viral hepatitis is prevalent. One important factor was the development of large reservoir of infection due to extensive schistosomiasis control programs that used intravenously administered tartar emetic since the 1960s.[52] Co-infection is known to cause earlier liver deterioration and more severe illness.[56]
Adult and larval worms migrate through the host's blood circulation avoiding the host's immune system. The worms have many tools that help in this evasion, including the tegument, antioxidant proteins, and defenses against host membrane attack complex (MAC).[57] The tegument coats the worm and acts as a physical barrier to host antibodies and complement. Host immune defenses are capable of producing superoxide, but these are counterattacked by antioxidant proteins produced by the parasite. Schistosomes have four superoxide dismutases, and levels of these proteins increase as the schistosome grows. Antioxidant pathways were first recognised as a chokepoints for schistosomes,[58] and later extended to other trematodes and cestodes. Targeting of this pathway with different inhibitors of the central antioxidant enzyme thioredoxin glutathione reductase (TGR) results in reduced viability of worms.[59] Decay accelerating factor (DAF) protein is present on the parasite tegument and protects host cells by blocking formation of MAC. In addition, schistosomes have six homologues of human CD59 which are strong inhibitors of MAC.[60]
The presence of S. mansoni is detected by microscopic examination of parasite eggs in stool. A staining method called Kato-Katz technique is used for stool examination. It involves methylene blue-stained cellophane soaked in glycerine or glass slides.[61] A costlier technique called formalin-ether concentration technique (FECT) is often used in combination with the direct faecal smear for higher accuracy. Serological and immunological tests are also available. Antibodies and antigens can be detected in the blood using ELISA to identify infection. Adult worm antigens can be detected by indirect haemagglutination assays (IHAs). Polymerase chain reaction (PCR) is also used for detecting the parasite DNA. Circulating cathodic antigen (CCA) in urine can be tested with lateral flow immune-chromatographic reagent strip and point-of-care (POC) tests.[62]
Egg detection and immunologic tests are not that sensitive.[63] Polymerase chain reaction (PCR) based testing is accurate and rapid.[63] They, however, are not frequency used in countries were the disease is common due to the cost of the equipment and the technical experience required to run them.[63] Using a microscope to detect eggs costs about US$0.40 per test well PCR is about $US7 per test as of 2019.[64] Loop-mediated isothermal amplification (LAMP) are being studied as they are lower cost.[63] LAMP testing is not commercially available as of 2019.[64]
The standard drug for S. mansoni infection is praziquantel at a dose of 40 mg/kg. Oxamniquine is also used.[65]
As of WHO report in 2016, 206.5 million people worldwide are having schistosomiasis due to different species of Schistosoma. More than 105 million people were given medical treatment.[1] S. mansoni is the major species causing an annual death of about 130,000.[66] It is endemic in 55 countries and most prevalent in Africa, the Middle East, the Caribbean, Brazil, Venezuela and Suriname.[67] About 80-85% of schistosomiasis is found in sub-Saharan Africa, where S. haematobium, S. intercalatum and S. mansoni are endemic. Approximately 393 million Africans are at risk of infection from S. mansoni, of which about 55 million are infected at any moment. Annual death due to S. mansoni is about 130,000.[68] The prevalence rate in different countries of Africa are: 73.9% in northern Ethiopia, 37.9% in western Ethiopia, 56% in Nigeria, 60.5% in Kenya, 64.3% in Tanzania, 19.8% in Ghana, and 53.8% in Côte d'Ivoire.[69] In Egypt, 60% of the population in the Northern and Eastern parts of the Nile Delta and only 6% in the Southern part are infected.[70]
S. mansoni is commonly found in places with poor sanitation. Because of the parasite's fecal-oral transmission, bodies of water that contain human waste can be infectious. Water that contains large populations of the intermediate host snail species is more likely to cause infection. Young children living in these areas are at greatest risk because of their tendency to swim and bathe in cercaria-infected waters longer than adults .[71] Anyone travelling to the areas described above, and who is exposed to contaminated water, is at risk of schistosomiasis.
The intermediate hosts Biomphalaria snails are estimated to originate in South America 95–110 million years ago. But the parasites Schistosoma originated in Asia. In Africa, the progenitor species evolved into modern S. mansoni and S. haematobium around 2–5 million years ago.[72][73]
A German physician Theodor Maximillian Bilharz was the first to discover the parasite in 1851, while working at Kasr el-Aini Hospital, a medical school in Cairo. Bilharz recovered them from autopsies of dead soldiers, and noticed two distinct parasites.[74] He described one of them as Distomum haematobium (now S. haematobium) in 1852,[75] but failed to identify the other. In one of his letters to his mentor Karl Theordor von Siebold, he mentioned some of the eggs were different in having terminal spines while some had lateral spines.[76] Terminal-spined eggs are unique to S. haematobium, while lateral spines are found only in S. mansoni. Bilharz also noted that the adult flukes were different in anatomy and number eggs they produced.[77] He introduced the terms bilharzia and bilharziasis for the name of the infection in 1856. A German zoologist David Friedrich Weinland corrected the genus name to Schistosoma in 1858; and introduced the disease name as schistosomiasis.[78]
The species distinction was first recognised by Patrick Manson at the London School of Hygiene & Tropical Medicine. Manson identified lateral-spined eggs in the faeces of a colonial officer earlier posted to the West Indies, and concluded that there were two species of Schistosoma.[79] An Italian-British physician Louis Westenra Sambon gave the new names Schistosomum haematobium and Schistosomum mansoni in 1907, the latter to honour Manson.[2] Sambon only gave partial description using a male worm. In 1908, a Brazilian physician Manuel Augusto Pirajá da Silva gave a complete description of male and female worms, including the lateral-spined eggs.[80] Pirajá da Silva obtained specimens from three necropsies and eggs from 20 stool examinations in Bahia.[81] He gave the name S. americanum.[82] The species identity was confirmed in 1907 by British parasitologist Robert Thomson Leiper,[79] identifying the specific snail host, and distinguishing the egg structure, thereby establishing the life cycle.[83]
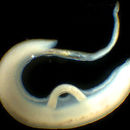
 A paired couple of Schistosoma mansoni.
A paired couple of Schistosoma mansoni. Schistosoma mansoni is a water-borne parasite of humans, and belongs to the group of blood flukes (Schistosoma). The adult lives in the blood vessels (mesenteric veins) near the human intestine. It causes intestinal schistosomiasis (similar to S. japonicum, S. mekongi, S. guineensis, and S. intercalatum). Clinical symptoms are caused by the eggs. As the leading cause of schistosomiasis in the world, it is the most prevalent parasite in humans. It is classified as a neglected tropical disease. As of 2021, the World Health Organization reports that 236.6 million people have schistosomiasis and most of it is due to S. mansoni. It is found in Africa, the Middle East, the Caribbean, Brazil, Venezuela and Suriname.
Unlike other flukes (trematodes) in which sexes are not separate (monoecious), schistosomes are unique in that adults are divided into males and females, thus, gonochoric. However, a permanent male-female pair, a condition called in copula, is required to become adults; for this, they are considered as hermaphrodites.
The life cycle of schistosomes includes two hosts: humans as definitive hosts, where the parasite undergoes sexual reproduction, and snails as intermediate hosts, where a series of asexual reproduction takes place. S. mansoni is transmitted through water, where freshwater snails of the genus Biomphalaria act as intermediate hosts. The larvae are able to live in water and infect the hosts by directly penetrating the skin. Prevention of infection is done by improved sanitation and killing the snails. Infection is treated with praziquantel.
S. mansoni was first noted by Theodor Maximillian Bilharz in Egypt in 1851, while discovering S. haematobium. Sir Patrick Manson identified it as unique species in 1902. Louis Westenra Sambon gave the name Schistosomum mansoni in 1907 in honour of Manson.
Schistosoma mansoni adalah parasit yang signifikan dari manusia, sebuah trematoda yaitu salah satu agen utama dari penyakit schistosomiasis. Para schistosomiasis disebabkan oleh Schistosoma mansoni adalah usus schistosomiasis.
Schistosomes adalah trematoda atipikal dalam tahap dewasa yang memiliki dua jenis kelamin (dioecious) dan terletak di pembuluh darah dari inang definitif. Sebagian besar trematoda lainnya adalah hermafrodit dan ditemukan di saluran usus atau organ, seperti hati. Siklus hidup schistosomes mencakup dua inang: inang definitif (yaitu manusia) di mana parasit mengalami reproduksi seksual, dan sejumlah siput tunggal menengah di mana ada beberapa tahap reproduksi aseksual. S. mansoni ini dinamai oleh Sir Patrick Manson, yang pertama kali diidentifikasi di Formosa (Taiwan).[2]
Schistosoma mansoni adalah parasit yang signifikan dari manusia, sebuah trematoda yaitu salah satu agen utama dari penyakit schistosomiasis. Para schistosomiasis disebabkan oleh Schistosoma mansoni adalah usus schistosomiasis.
Schistosomes adalah trematoda atipikal dalam tahap dewasa yang memiliki dua jenis kelamin (dioecious) dan terletak di pembuluh darah dari inang definitif. Sebagian besar trematoda lainnya adalah hermafrodit dan ditemukan di saluran usus atau organ, seperti hati. Siklus hidup schistosomes mencakup dua inang: inang definitif (yaitu manusia) di mana parasit mengalami reproduksi seksual, dan sejumlah siput tunggal menengah di mana ada beberapa tahap reproduksi aseksual. S. mansoni ini dinamai oleh Sir Patrick Manson, yang pertama kali diidentifikasi di Formosa (Taiwan).
Schistosoma mansoni is een tropische parasitaire worm uit het geslacht Schistosoma, die de ziekte schistosomiasis veroorzaakt. Jaarlijks raken circa 210 miljoen mensen ermee besmet. In juli 2009 werd bekend dat onderzoekers van het Leids Universitair Medisch Centrum de genetische code van de worm ontrafeld hebben, wat op termijn een bijdrage moet leveren aan een medicijn.[1]
Schistosoma mansoni heeft een ingewikkelde levenscyclus. Uit wormeitjes ontwikkelen zich in zoet water een larvestadium (miracidia), die zich vrij in het water voortbewegen door middel van trilhaartjes. Deze larven zijn ofwel mannelijk, ofwel vrouwelijk. Ze dringen een tussengastheer (een zoetwaterslak) binnen.
In de slak ondergaan de larven een aantal ontwikkelingsstadia. De miracidia verliezen hun trilharen en veranderen in een moedersporocyst: een soort zak waarin de larve zich ongeslachtelijk vermeerdert. Iedere moedersporocyst produceert meerdere dochtersporocysten, die op hun beurt weer een ander type larve (cercaria) in grote hoeveelheden voortbrengen. Ieder miracidium brengt zodoende honderdduizenden cercaria voort. Ook hiervan zijn er mannelijke en vrouwelijke exemplaren.
De cercaria verlaten hun tussengastheer en zwemmen naar hun tweede gastheer. Als mensen in besmet water baden kan de parasiet in dit larvestadium ongemerkt door de huid het lichaam binnendringen. Eenmaal in de bloedbaan reizen de piepkleine cercaria naar de lever en groeien uit tot wormen van ongeveer een centimeter. Mannetjes en vrouwtjes gaan vervolgens in paren naar de bloedvaten rond de darmen om eitjes te leggen. Ongeveer twee maanden na een besmetting zijn de microscopisch kleine eitjes aantoonbaar in de ontlasting. Hiermee is de cyclus rond. Dit proces, van eitje tot eileggende worm duurt 25 tot 30 dagen.
De levensduur van de Schistosoma mansoni kan meer dan twintig jaar bedragen, maar is meestal ongeveer vijf jaar.
Cercaria larve van Schistosoma mansoni
eitje van Schistosoma mansoni
Schistosoma mansoni is een tropische parasitaire worm uit het geslacht Schistosoma, die de ziekte schistosomiasis veroorzaakt. Jaarlijks raken circa 210 miljoen mensen ermee besmet. In juli 2009 werd bekend dat onderzoekers van het Leids Universitair Medisch Centrum de genetische code van de worm ontrafeld hebben, wat op termijn een bijdrage moet leveren aan een medicijn.
Przywra żylna (Schistosoma mansoni) – dwużywicielska pasożytnicza przywra, wywołująca schistosomatozę. Jej żywicielem pośrednim jest wodny ślimak, a ostatecznym kręgowiec – człowiek. Pasożytuje w żyłach krezki jelitowej[2].
Dorosłe przywry są rozdzielnopłciowe, wykazują dymorfizm płciowy. Osiągają 10–15 mm (samce) lub 14–20 mm (samice)[3] i są najmniejszymi przedstawicielami rodzaju Schistosoma[4]. Ich ciało jest wydłużone, samce są szersze od samic, posiadają kanał gynekoforyczny na brzusznej stronie ciała, w którym umieszczona jest samica podczas stałej kopulacji. U samca występuje 6–9 jąder[5], a u samic pojedynczy jajnik położony z przodu ciała[4]. Posiadają dwie przyssawki – gębową i brzuszną oraz kolce tegumentalne, służące do przytwierdzania się do ciała żywiciela[6]. Powierzchnia ciała samca pokryta jest niewielkimi wyrostkami, u samicy występują one jedynie na przednim i końcowym odcinku ciała poza obszarem, który znajduje się w kanale gynekoforycznym[5].
Ze względu na rozdzielnopłciowość S. mansoni w żywicielu ostatecznym konieczne jest występowanie zarówno samca jak i samicy, aby mógł zajść proces rozmnażania. Samce w zarażeniu jednopłciowym przechodzą pełen rozwój, natomiast samice dojrzewają tylko w obecności samca. Proces dojrzewania samicy może ulec regresji w wyniku przeniesienia jej do nowego żywiciela, w którego organizmie nie występują samce[6]. Do osiągnięcia dojrzałości przez samicę nie jest wymagane zapłodnienie, lecz długotrwałe przebywanie w kanale gynekoforycznym. Samce w zarażeniu jednopłciowym mogą wykształcić cechy charakterystyczne dla samic. Osobniki oprócz par złożonych z samca i samicy mogą także tworzyć pary homoseksualne, a także grupy złożone z wielu osobników[5].
Osobniki S. mansoni mogą krzyżować się także z osobnikami należącymi do innych gatunków. W wyniku tego procesu powstają hybrydy międzygatunkowe, które osiągają większe rozmiary, składają większą liczbę jaj, a proces osiągania dojrzałości płciowej trwa u nich krócej[6].
Mimo życia w środowisku bogatym w tlen (naczynia krwionośne) metabolizm dorosłych przywr żylnych jest w większości beztlenowy. Tlen jest niezbędny do utwardzania jaj[6].
S. mansoni żyją w układzie krwionośnym żywiciela ostatecznego, dlatego żywią się krwią. Energię uzyskują głównie z procesu utleniania glukozy. Przywra w godzinę jest w stanie przyjąć ilość pokarmu odpowiadającą 15–20% masy jej ciała. W tym czasie para osobników może zjeść nawet 350 000 erytrocytów. Trawienie hemoglobiny zachodzi w przewodzie pokarmowym przywry[6][7].
Jajo S. mansoni posiada kolec w dolnej części, który ułatwia mu przejście przez ściany naczyń krwionośnych[6]. Dorosła samica składa od 100 do 300 jaj w ciągu dnia. Optymalna temperatura potrzebna do wyklucia się z jaj osobników potomnych wynosi 28°C. W temperaturze 4°C oraz 37°C proces ten zostaje prawie całkowicie zahamowany[5].
Żywicielem pośrednim przywry są ślimaki z rodzaju Biomphalaria, m.in. B. pfeifferi, B. camerunensis, B. sudanica i B. alexandrica, a w Ameryce Południowej ślimaki należące do rodzaju Tropicorbis. Żywicielem ostatecznym może zostać człowiek, gryzonie, oposy, a także niektóre małpy (koczkodany i pawiany)[4][6].
Zapłodnione samice przywr składają jaja w naczyniach krwionośnych żywiciela ostatecznego. Jaja migrują przez tkanki do światła jelita. Wraz z fekaliami dostają się do środowiska wodnego. Z jaj pod wpływem optymalnych warunków świetlnych i cieplnych wylęgają się miracidia, które wnikają do organizmu żywiciela pośredniego – słodkowodnego ślimaka z rodzaju Biomphalaria. Penetrują tkanki ślimaka, a następnie ulegają przekształceniu w kolejne stadia rozwojowe – dwie generacje sporocyst. Kolejne stadium rozwojowe stanowią ruchliwe cerkarie powstające w wyniku rozmnażania bezpłciowego w gruczole wątrobowym ślimaka. Opuszczają one ciało ślimaka w poszukiwaniu żywiciela ostatecznego – kręgowca. Proces ten odbywa się z 30-dniowymi przerwami, w czasie których pozostałe zarodki cerkarii ulegają degeneracji, dzięki czemu możliwe jest powstanie kolejnej generacji sporocyst potomnych. Wychodzenie cerkarii z ciała ślimaka odbywa się według określonego rytmu, który rozpoczyna się rano, osiąga maksimum około południa, a kończy wieczorem. Uwolnione ze ślimaka cerkarie dostają się do organizmu żywiciela ostatecznego poprzez penetrację skóry i przekształcają się w schistosomule wędrujące przez naczynia krwionośne do serca, płuc, z powrotem do serca, a następnie do układu wrotnego wątroby, w którym dojrzewają i stają się dorosłymi osobnikami. Dojrzałe przywry łączą się w pary, kopulują i wędrują do żył krezkowych, gdzie składają jaja[2][6][7].
Jaja S. mansoni powodują zapalenie błony śluzowej jelita grubego, czego wynikiem są owrzodzenia i brodawczaki, a także zmiany nowotworowe. Jaja wraz z krwią mogą dostać się do innych narządów, np. wątroby, gdzie w wyniku zablokowania światła naczyń żylnych powodują powiększenie tego narządu, a także śledziony. Prowadzi to również do powstania obrzęków, wodobrzusza i żółtaczki. Jaja mogą trafić także do płuc, mózgu, a nawet oczu[4][6].
W celu wykrycia obecności jaj pasożyta wykonuje się badanie kału, badania endoskopowe odbytnicy, biopsję zmienionej tkanki, a także analizę serologiczną[4].
W leczeniu stosuje się leki przeciwpasożytnicze – oksamnichinę i prazykwantel. Aby uniknąć zarażenia pasożytem, należy pić tylko przegotowaną wodę i unikać kąpieli w otwartych zbiornikach wodnych[4].
W Afryce występowanie S. mansoni pokrywa się z rozprzestrzenieniem S. haematobium. W Ameryce Południowej, gdzie została zawleczona w XVI–XVII wieku, występuje na terenie Brazylii, Wenezueli i na Dominikanie. Jest obecna także w Azji na Półwyspie Arabskim[4][6].
Przywra żylna (Schistosoma mansoni) – dwużywicielska pasożytnicza przywra, wywołująca schistosomatozę. Jej żywicielem pośrednim jest wodny ślimak, a ostatecznym kręgowiec – człowiek. Pasożytuje w żyłach krezki jelitowej.
Schistosoma mansoni är en plattmaskart som ingår i släktet Schistosoma, och familjen Schistosomatidae.[1] Plattmasken är en parasit som kan ge upphov till den kroniska sjukdomen Schistosomiasis, även kallat snäckfeber eller bilharzia, som kan vara mycket besvärlig. Parasiten finns framförallt i Afrika men även i andra världsdelar där ökad utbredning sker.
Det finns ett 20-tal arter inom schistosomasläktet[1] varav fem av dem kan orsaka sjukdomen snäckfeber. Schistosoma mansoni, Schistosoma japonicum och Schistosoma haematobium är de arter som i störst utsträckning orsakar sjukdomen medan de andra två arterna, Schistosoma intercalatum[2] samt Schistosoma mekongi, är mindre vanliga, dock med liknande symptom som S. japonicum och S. haematobium.[3]
Schistosoma mansoni har en trattformad sugmun i ena änden och ett ventralt sugorgan i direktkontakt med tarmen.[4] Maskarna har en gul eller gulbrun färgton och könen har olika kroppsbyggnad.[5] Hanen är cirka 10 mm lång och 1 mm bred och vit till färgen. Hanen har 6–9 testiklar och de genitala porerna sitter i den bakre änden nära det ventrala sugorganet. Honan är smalare och något längre (10–14 mm) och har en enda äggstock belägen i främre delen av kroppen. Livmodern kan ha olika storlek beroende på var någonstans äggstocken sitter i förhållande till den genitala poren. Sitter äggstocken nära den genitala poren är livmodern kortare än om den sitter längre ifrån.[4]
Schistosoma mansoni sägs härstamma från Asien och skall ha spridits till Afrika. Parasiten framträdde förmodligen efter att sötvattensnäckor av släktet Biomphalaria spred sig över Atlanten från neotropikerna till just Afrika. Detta skedde, enligt studier som gjorts på afrikanska fossiler, för 2–5 miljoner år sedan.[6]
Parasitpopulationen utvecklades framgångsrikt i Afrika och spreds med slavhandeln vidare till både Europa och Amerika.[6] I Egypten har Schistosoma mansoni nästan helt ersatt släktingen Schistosoma haematobium i nildeltat och därifrån spridits ut över hela landet. I Senegal har byggandet av dammar och vattenbarriärer lett till en epidemi och i Brasilien har parasiten lokalt blivit ett problem.[6] Trots den stora utbredningen i världen har parasiten på de karibiska öarna blivit utkonkurrerad av andra vattenlevande organismer och påverkats av miljöförändringar.[6]
Det är i plattmaskens larvstadium som den ”attackerar” människor när de badar i sötvatten där arten förekommer. Larven penetrerar huden i vattnet och i kroppen vidareutvecklas den till en vuxen schistosom som lever i blodkärlen. Honorna producerar därefter ägg som sprids via avföring eller urin medan andra fastnar i kroppsvävnaden.[7]
Vuxna Schistosoma mansonis är ungefär 10 mm långa. Honan och hanen formar tillsammans ett reproduktivt par i tarmen hos värden där honan ligger som i en liten ficka hos hanen. Ett par kan ligga i flera år hos denna värd och honan producerar ofta flera tusentals ägg under denna period.[8]
De nyproducerade äggen släpps genast ut genom tarmväggarna och cirkulerar sedan runt i blodet. Vissa av äggen fastnar i levern och i andra inre organ och skapar på så vis många av de symtom och besvär som till slut kommer att leda till sjukdomen schistosomiasis. De ägg som inte fastnar i organen följer med avföringen samt urin och om de i samband med detta kommer i kontakt med sötvatten kommer äggen att mogna och kläckas. När de kläcks kommer de gå över till sitt ”frilevande” larvstadium som kallas miracidium.[8]
Miracidielarver simmar runt i sötvatten med hjälp av cilier som de har på sin kropp. De är kortlivade och lever enbart i en dag. För att miracidier ska kunna överleva krävs det därför att det snabbt hittar och infekterar en annan värd, nämligen sötvattensnäckor av släktet Biomphalaria. Inuti snäckorna omvandlas miracidier till sporocyster. I sporocyststadiet produceras cercarielarver genom asexuell reproduktion. På så vis kan en miracidie (i sitt sporocyststadium) producera flera tusentals genetiskt identiska cercarier i mellanvärden.[8] Biomphalariasnäckorna börjar sedan, 3–4 veckor efter att de blivit infekterade av en miracidie, att släppa ut cercarielarver i sötvattnet där de kan simma runt fritt. Även detta stadium är kortvarigt (lever i cirka en dag). För att överleva måste cercarielarven denna gång hitta en människa som blir larvens slutvärd och det är på så vis parasiten infekterar människor.
Det tar ca en månad för en cercarielarv att inuti människan utvecklas till en vuxen Schistosoma mansoni, emigrera till blodkärlen runt tarmen, hitta en partner och börja producera ägg.[8]
Schistosomiasis är en kronisk sjukdom som orsakar stora hälsoproblem och i dagsläget är omkring 83 miljoner människor infekterade med Schistosoma mansoni.[9]
Några dagar efter att man har blivit infekterad kan det uppstå klåda och utslag på huden men de allra flesta känner inte av dessa symptom. Efter 1-2 månader drabbas man av feber, hosta och muskelsmärtor. De ägg som inte passerar ut ur kroppen med avföringen/urinen lägger sig i urinblåsan eller i tarmväggen och kan där orsaka inflammation och ärrbildning. Det är alltså dessa ägg snarare än plattmaskarna som orsakar symptomen för schistosomiasis.[10]
Upprepade infektioner kan leda till undernäring, blodbrist och inlärningssvårigheter hos barn. Även levern, mjälten, lungorna, urinblåsan och tarmarna kan skadas av parasiten om man bär på den under flera år. Utan behandling kan schistosomiasis bli kroniskt vilket förhöjer risken för cancer i urinblåsan med symptom såsom; buksmärta, förstorad lever, svårigheter med att urinera samt blod i avföringen/urinen. Vid enstaka fall kan äggen ta sig till hjärnan och ryggmärgen och det kan yttra sig i kramper, förlamning eller inflammation i ryggmärgen.[10]
Jordbrukets utveckling i många länder medför krav på utökade vattensystem och dessa har lett till en ökad spridning av framförallt Schistosoma mansoni.[11] Global uppvärmning kan komma att leda till en spridning av parasitens värdar, Biomphalariasnäckor.[6]
Det finns medicinering för schistosomiasis, Praziquantel, som ska intas i 1-2 dagar [12] samt läkemedelet Meclonazepam som har en scistodomicidal aktivitet gentemot Schistosoma mansoni [13]
Det krävs att denna behandling påbörjas i tid för att inte sjukdomen ska bli kronisk. God hygien på toaletter samt bekämpning mot Biomphalariasnäckorna minskar riskerna för sjukdomen. För att undvika att bli infekterad bör man ej bada eller dricka vatten i floder och sjöar i riskzonerna.[14] Det forskas även på att framställa ett vaccin som gör det omöjligt för parasiten att slutföra sin livscykel i människan.[6]
Schistosoma mansoni är en plattmaskart som ingår i släktet Schistosoma, och familjen Schistosomatidae. Plattmasken är en parasit som kan ge upphov till den kroniska sjukdomen Schistosomiasis, även kallat snäckfeber eller bilharzia, som kan vara mycket besvärlig. Parasiten finns framförallt i Afrika men även i andra världsdelar där ökad utbredning sker.
Schistosoma mansoni — важливий патоген людини, один з головних збудників шистосомозу. Цей організм нетиповий серед трематод тим, що в дорослому стані має дві статі та мешкає у кровоносних судинах хазяїна, тоді як решта траматод — гермафродити, що мешкають у кишечнику або органах, таких як печінка. Життєвий цикл цього організму складається з двох стадій: головного хазяїна (наприклад, людини), де він проходить статеве розмноження, та існування у равликах, як безстатевий організм.
Schistosoma mansoni — важливий патоген людини, один з головних збудників шистосомозу. Цей організм нетиповий серед трематод тим, що в дорослому стані має дві статі та мешкає у кровоносних судинах хазяїна, тоді як решта траматод — гермафродити, що мешкають у кишечнику або органах, таких як печінка. Життєвий цикл цього організму складається з двох стадій: головного хазяїна (наприклад, людини), де він проходить статеве розмноження, та існування у равликах, як безстатевий організм.

マンソン住血吸虫(マンソンじゅうけつきゅうちゅう、学名:Schistosoma mansoni)は、住血吸虫科住血吸虫属に属する吸虫の1種。Biomphalaria属の淡水巻貝を中間宿主、ヒト、サル、ネズミを終宿主とし、門脈に寄生して腸管住血吸虫症を引き起こす。名前はスコットランド出身の医師パトリック・マンソン(英語版)が台湾でこの吸虫を見出したことに因んで付けられた。
治療にはオキサムニキンが用いられる。ただし、アフリカ株は南米株と比べてオキサムニキンに対し耐性を有することが報告されている[2]。
成虫は吸虫としては珍しく、細長い線虫様の形態をしている。 雌雄異体で、終宿主には雄が抱雌管で雌を抱えた状態で寄生する。 雄は白く、体長1cm程度、太さは1mm程度である。 雌は灰色で、体長はやや長く、雄と比べるとかなり細い。雌の重量のほとんどが生殖器で占められている。 消化管は口吸盤に始まり、食道が2分岐して再び1つにまとまったのち、行き止まりになっていて肛門はない。 虫卵は黄褐色で、側方部に刺状の突起がある。虫卵は小蓋を持たない。
虫卵は糞便とともに排出され、水中で成熟ミラシジウムが孵化する。
ヒラマキガイ科Biomphalaria属に属する巻貝を中間宿主としており、ミラシジウムはこの貝に出会うと体表から組織内へと侵入しスポロシストへと変態する。母スポロシスト、娘スポロシストの2世代を過ごした後、数千のセルカリアが生じる。なおベネズエラではアフリカマイマイも中間宿主となりうることが報告されている。 [3]
セルカリアは日中に貝から水中へと泳ぎ出し、終宿主と出会うと皮膚から体内へと侵入する。 このときセルカリアは皮膚を這い毛嚢などの好適部位を探し出して、タンパク質分解酵素を分泌して頭部を皮膚組織へと侵入させる。 頭部はシストソミューラに変態し、皮膚に数日留まったあと血流に乗って肺へと移動する。 成虫は肝臓の類洞へとたどりつくと、つがいをつくり血流に逆らって腸間膜静脈に移動して産卵を始める。 [4][5]
雌の成虫は雄の抱雌管にはさまれている。 雄は雌がいてもいなくても正常に成熟できるが、それでも行動や生理には差が生じることが知られている。 一方、雌は雄がいないと成熟することができず、生殖器が発達しない。
マンソン住血吸虫の成虫は、大腸や盲腸の周囲の下腸間膜静脈に寄生し、 一日あたりおよそ300個の虫卵を毛細血管内皮に産み付ける [6] 虫卵は腸管内に入り込み、糞便と共に宿主体外へと排出される。
ゲノムサイズはおよそ270 Mbpで、GC含量は34%である。ゲノムの約4割を反復配列が占めている。 染色体は8対で、そのうち1対が性染色体である。 受精の時点で染色体により性が決定され、哺乳類とは逆に、同型で雄(ZZ)、異型で雌(ZW)となる。 [7] 2009年にゲノムの概要が解読され、タンパク質分解酵素が多様化していることと、脂肪酸やステロールの生合成ができないことが特徴として示された。 [8][9]
住血吸虫のなかでも広範囲に分布しており、アフリカ、中東、カリブ海沿岸、南米など世界でおよそ8000万人が感染しているとされる。[10]
マンソン住血吸虫(マンソンじゅうけつきゅうちゅう、学名:Schistosoma mansoni)は、住血吸虫科住血吸虫属に属する吸虫の1種。Biomphalaria属の淡水巻貝を中間宿主、ヒト、サル、ネズミを終宿主とし、門脈に寄生して腸管住血吸虫症を引き起こす。名前はスコットランド出身の医師パトリック・マンソン(英語版)が台湾でこの吸虫を見出したことに因んで付けられた。
治療にはオキサムニキンが用いられる。ただし、アフリカ株は南米株と比べてオキサムニキンに対し耐性を有することが報告されている。











