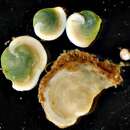
Live individuals of Archaias angulatus are easily recognizable by the bright “grass-green” coloration of their cytoplasm (Hallock and Peebles 1993). This distinctive coloration results from the mutualistic association of the living foraminiferan with intracellular chlorophycean algae. Transmission electron microscopy studies show the symbiotic algae to be distributed throughout the cytoplasm of the foraminiferal host cell, from the inner to outermost chambers (Müller‐Merz and Lee 1976). Within individual chambers, the symbiotic algae are observed to cluster under the thin chamber walls of the foraminiferal shell, “like plants in a greenhouse” (Lee et al. 1974, Müller‐Merz and Lee 1976). Symbiotic algae lying within the host cytoplasm are enclosed by a host-derived symbiosome membrane (Lee 2002). The algal cells are spherical in shape, range in size from 10- to 14-µm in diameter, and lack the flagella and thick walls characteristic of free-living algae growing in axenic culture (Lee and Bock 1976, Lee et al. 1974).
The algal endosymbionts associated with Archaias angulatus can be cultured independently of the foraminiferal host cell (Lee et al. 1974, Lee and Zucker 1969). Lee and Zucker (1969) were the first researchers to identify the algal endosymbionts isolated from A. angulatus as chlorophycean algae. These chlorophycean algae were subsequently described as a new species, Chlamydomonas hedleyi, by Lee et al. (1974). Molecular phylogenetic analyses demonstrate that Chlamydomonas hedleyi is a member of the more inclusive “Moewusii” clade of chlorophycean algae (Pröschold et al. 2001), and is closely related to free-living species marine and freshwater species, including C. raudensis, a psychrophilic green alga isolated from 17 m below the permanent ice cover of Lake Bonney, Antarctica (Gerloff-Elias et al. 2005, Klochkova et al. 2008, Pocock et al. 2004). In another molecular phylogenetic study, Chlamydomonas hedleyi branches as the basal lineage in a monophyletic clade of chlorophycean endosymbionts associated with other species of Soritacean foraminiferans (Pawlowski et al. 2001). More than one phylotype of chlorophyceans may be present in an individual, population, or species. Some of the chlorophycean endosymbionts associated with Archaias angulatus are more closely related to the chlorophycean endosymbionts associated with living species of Laevipeneroplis bradyi and L. proteus, than to Chlamydomonas hedleyi Lee, Crockett and Stone, 1974 (Pawlowski et al. 2001).
The endosymbiotic relationship between the foraminiferal host and the chlorophycean algae is an example of “persistent obligatory acquired phototrophy” sensu Stoecker et al. (2009), an intermediate type of acquired phototrophy in which the algal endosymbiont is always present in living Archaias angulatus; however, the foraminiferal host can not grow in the absence its endosymbionts.
Archaias angulatus benefits from its mutualistic association with chlorophycean algae in several different ways. First, the products of photosynthesis (sugars and energy byproducts) can be transferred from the algae to the host cell. Lee et al. (1974) estimated that as much as 57% of the carbon fixed by Chlamydomonas hedleyi is released in the form of the sugar mannitol, while 43% of the carbon fixed is retained by the algae. Starch grains have been observed lying free in the cytoplasm of the foraminiferal host. It has also been suggested that the foraminiferan host received energy products in the form of ATP produced during photosynthesis of the algal endosymbionts, although this process has not been investigated in A. angulatus. Second, calcification of the foraminiferal shell is enhanced by algal photosynthesis. Duguay and Taylor (1978) observed light-enhanced calcification (45C uptake) in A. angulatus to be directly proportional to the photosynthetic rate (14C uptake) of the symbiont C. hedleyi, and light flux, while individuals of Archaias angulatus treated with the herbicide DCMU (1-100 µM) exhibited inhibition of photosynthesis and light-enhanced calcification (Duguay and Taylor 1978). Lastly, the algae may represent a potential food source for the host foraminiferan. Archaias angulatus has been observed to digest its endosymbionts when starved (Hallock and Peebles 1993), which may contribute to its ability to survive a few weeks without feeding (Hallock in Walker et al. 2011).
The algal endosymbionts of Archaias angulatus are also believed to benefit from their symbiotic association with the host foraminiferal cell (Hallock 2003, Lee 2006). The intracellular milieu of the host foraminiferan may represent an important source of nitrogen in the oligotrophic waters of coral reef and associated habitats. In other endosymbiotic associations, the host transfers a portion of its metabolic nitrogenous waste to the symbiotic algae (Cook 1972, Nowack and Melkonian 2010). Lee et al. (1974) found urea to be the best nitrogen source for C. hedleyi, although the preferred source of nitrogen in most species of Chlamydomonas is ammonium (Fernández et al. 2009),Like other species of Chlamydomonas, C. hedleyi grown in axenic culture exhibits no specific vitamin requirements; however, thiamine has been shown to be a growth inducer, and correspondingly, growth in C. hedleyi is increased three-fold in the presence of thiamine (Croft et al. 2006, Lee 2006, Lee et al. 1974).
Archaias angulatus has a planispirally-coiled, flabelliform test. The overall test morphology is biconvex and lenticular in shape with a keeled periphery. The mode of coiling is initially involute with each new whorl covering previous whorls. In the terminal flabelliform stage, coiling becomes advolute (partially evolute).
Archaias angulatus has a porcelaneous wall structure, in which the primary wall layer is composed of a three-dimensional random array of lath-like crystals of high-magnesium calcite (Towe and Cifelli 1967). The magnesium content of the calcite crystals ranges from 14-16 mol% (Blackmon and Todd 1959). The primary wall layer is sandwiched between a thin outer veneer of flattened two-dimensionally oriented crystals that coats the surface of the chambers, and an inner basal imperforate layer from which the endoskeletal elements are formed.
Each individual chamber is shaped like a coiled strip that bifurcates into two three-dimensional spirals, the alar prolongations, each of which terminates at the axis of coiling (Hottinger 2006: Fig. 7A, B). The chambers are greater in breadth than height, and are of relatively uniform height throughout the length of the chamber. Archaias angulatus, like other milioline foraminiferans, has an accretionary mode of growth, in which new chambers are formed episodically throughout the lifespan of an individual cell. Each new chamber that is formed is cemented on to the previous chamber in a separate growth event. There is minimal overlap between successive chambers; the sutures, or zones of contact between the successive chambers, are slightly depressed and spiral in shape.
The lateral surfaces of each chamber are penetrated by numerous, large, rounded pseudopores that are haphazardly distributed over most of the distal chamber face (Lynts and Pfister 1967). Pseudopores, also called pits, pit-like pores or punctations (Hottinger 2001), are blind pores that penetrate the shell surface, but do not perforate the test wall. The pseudopores are approximately 5-6 µm in diameter, and occur at a density of less than 1 pit per 100 µm2 of chamber surface area (Cottey and Hallock 1988, Richardson 2000). An imperforate band is present along the base (proximal edge) of the chamber face adjacent to the suture (Crapon de Caprona d'Ersu and Benier 1985, Lévy 1977, Richardson 2000).
Each chamber possesses multiple apertures that are arranged in offset rows parallel to the long axis of the apertural face (Lévy 1977). Two rows of apertures are present on each of the alar prolongations; these two sets of apertural rows merge into four or more rows of apertures on the primary apertural face (Lévy 1977). The apertures are circular, subcircular to oval in shape and are bordered by a rounded, elevated, imperforate rim (peristome of Hottinger (2006)). Oval-shaped apertures are oblong, with the long axis of the aperture parallel to the long axis of the apertural face. The apertural face is ornamented with a network of reticulate ridges and depressions that impart a spongy texture to the apertural face.
Endoskeletal elements present in the interior of each chamber include rounded pillars and socculi. Pillars are cylindrical or columnar structures that extend from the chamber floor to the chamber roof (Gudmundsson 1994, Hamaoui and Brun 1974, Hottinger 2001, 2006, Seiglie et al. 1977, Smout and Eames 1958). Pillars (interseptal pillars or pilintradermal plates) are free-standing structures within the chamber lumen that are not connected laterally to the chamber walls. There are three-to-four rows of pillars present in terminal chambers of adult specimens of Archaias angulatus. The pillars arise from low raised ridges (socculi) that form on the floor of the chamber lumen from the basal (inner wall) layer of calcite that is deposited over the apertural face of the previous chamber (Hottinger 2001, 2006, Seiglie et al. 1977). Socculi may occur as either parallel ridges or an interconnected network of ridges. The apertures of the previous chamber (intercameral foramen) are located in depressions (sulci) between the socculi (Seiglie et al. 1977). Pillars occur at regular intervals within the chamber lumen, spaced at 50-µm intervals, and are visible in live specimens or wetted dead tests viewed in transmitted light (Cottey and Hallock 1988).
Living individuals of Archaias angulatus possess a “grass-green” coloration due to the presence of unicellular chlorophyte endosymbionts in their cytoplasm (Hallock and Peebles 1993).
Both total and partial SSU ribosomal DNA sequences have been published for six specimens of Archaias angulatus collected from the Florida Keys, Puerto Rico, and Brazil (Holzmann 2010b, Holzmann et al. 2001).
The following list gives the accession numbers for the six sequences deposited in GenBank (Benson et al. 2013). GenBank accession number AJ404302 represents the total 18S rRNA gene sequence for specimen 879 of Archaias angulatus collected from Long Key, Florida, USA. GenBank accession number FM877688 represents the total 18S rRNA gene sequence for specimen 6668 of Archaias angulatus collected from Maracajau Reef, Brazil. GenBank accession number AJ278046 represents a partial 18S rRNA gene sequence for specimen 676 of Archaias angulatus collected from Long Key, Florida, USA. GenBank accession number AJ278047 represents a partial 18S rRNA gene sequence for specimen 72 of Archaias angulatus collected from Mayagüez, Puerto Rico. GenBank accession number FM877690 represents a partial 18S rRNA gene sequence for specimen 71: clone 1 of Archaias angulatus collected from Mayagüez, Puerto Rico. GenBank accession number Z69603 represents a partial 18S rRNA gene sequence for Archaias angulatus collected from Mayagüez, Puerto Rico.
DNA barcodes for foraminiferans have been developed from six hypervariable expansion segments of the 18S rDNA gene: 37f, 41f, 43f, 45e, 47f, and 49e (Pawlowski and Lecroq 2010). The length of sequence varies from 1000-1200 nucleotides, depending on the species (Holzmann 2010a). Barcode sequences have been published for specimens 6668, 676, 71, 72, and 879 listed above, and are available from the foramBarcoding: Molecular Database of Foraminifera website (Holzmann 2010b).
Archaias angulatus is a microherbivore that obtains most of its carbon from grazing on marine bacteria and microalgae (Duguay and Taylor 1978, Lee and Bock 1976). Archaias angulatus obtains twice as much carbon through feeding, as it does from the transfer of photosynthetic products from its algal endosymbionts (Lee and Zucker 1969). As much as 57% of the carbon fixed by the algal symbiont Chlamydomonas hedleyi may be released to the host foraminiferal cell in the form of mannitol (or glucose) (Lee et al. 1974). In laboratory experiments, A. angulatus feeds primarily on bacteria and diatoms, eating large quantities of Cylindrotheca closterium, Navicula diversistriata, and Nitzschia acicularis, and smaller quantities of Nitzschia sp. and an unidentified Nannochloris-like chlorophyte (Lee and Zucker 1969).
Feeding rates in A. angulatus, determined using radioactively labeled (14C) microalgae, show that the foraminiferan feeds selectively on different prey items (Lee and Bock 1976). The highest feeding rates were observed when specimens of A. angulatus were fed the microalgal species Amphora coffeaeformis (0.97 g C/g organic weight of foram/0.25 day), Mastoglea sp. (0.9 g C/g organic weight of foram/0.25 day), Coconeis placentula (0.8 g C/g organic weight of foram/0.25 day), and Chlamydomonas hedleyi (0.7 g C/g organic weight of foram/0.25 day) (Lee and Bock 1976). Lower feeding rates were observed when the foraminiferan was fed Achnanthes haukiana (0.33 g C/g organic weight of foram/0.25 day), Phaeodactylum tricornutum (0.1 g C/g organic weight of foram/0.25 day), Nitzschia acicularis (0.1 g C/g organic weight of foram/0.25 day), and Dunaliella salina (0.1 g C/g organic weight of foram/0.25 day) (Lee and Bock 1976). Little overlap was observed between the diet of Archaias angulatus, and that of the epiphytic foraminifera Sorites dominicensis, except for Mastoglea sp., which both species consumed in large quantities (Lee and Bock 1976)
Transmission electron microscopy of A. angulatus cells show digestive vacuoles to be restricted to the outer the outer chambers of the shell, a feature that has been observed in other foraminiferal species (Müller‐Merz and Lee 1976). Archaias angulatus digests its chlorophycean symbionts when starved (Hallock and Peebles 1993), and can survive a few weeks without feeding (Hallock in Walker et al. 2011).
The only recorded predator of Archaias angulatus is the ornate blue crab, Callinectes ornatus (Haefner Jr 1990). Shells of A. angulatus were present in 9% of foreguts of Callinectes ornatus crabs collected from Thalassia meadows in Mullet Bay, Bermuda (Haefner Jr 1990). The most frequently occurring prey items found in crab guts were button snails (Modulus modulus) and Cerithium lutosum gastropods, other marine organisms associated with Thalassia blades (Vermeij 1978). The crabs appear to ingest pieces of Thalassia blade while capturing their snail prey, so it seems likely that the foraminiferan shells were incidentally ingested along with fragments of the seagrass blades.
Nautilus angulatus was first described and figured by Fichtel and Moll (1798: p. 113, pl. 22, figs. a-e). Rögl and Hansen (1984) designated a lectotype (MI-568), and two paralectotypes (MI-569, MI-570) from the three original syntypes, and redescribed and refigured all of the type material of Nautilus angulatus Fichtel and Moll, 1798 (p. 69, pl. 27, figs. 3-4, pl. 28, pl. 28, figs. 2-6).
Two other species of Nautilus described by Fichtel and Moll (1798), N. orbiculus and N. aduncus, represent different developmental stages of angulatus, and have since been synonymized with Nautilus angulatus (Crapon de Caprona d'Ersu and Benier 1985, Cushman 1946, Lévy 1977, Rögl and Hansen 1984). Nautilus orbiculus Fichtel and Moll (1798: p. 112, pl. 21, figs. a-e) represents a juvenile stage of N. angulatus; the holotype of N. orbiculus (MI-567) has been redescribed and refigured by Rögl and Hansen (1984: p. 69, pl. 27, figs. 1, 2). Nautilus aduncus Fichtel and Moll (1798: p. 115, pl. 23, figs. a-e) represents the adult, flabelliform stage of N. angulatus; the lectotype (MI-571) and paralectotype of N. aduncus have been redescribed and refigured by Rögl and Hansen (1984: p. 70, pl. 27, figs. 5-6, pl. 29, figs. 1-4).
The type specimens of Archaias angulatus, A. orbiculus, and A. aduncus are housed in the collections of the Department of Geology and Paleontology, Natural History Museum, Vienna, Austria.
In Conchyliologie systématique et classification méthodique des coquilles, De Montfort (1808) designated the three Fichtel and Moll (1798) species N. angulatus, N. aduncus, and N. orbiculus, as the type species for three new genera, Archaias, Helenis, and Ilotes, respectively. De Montfort (1808) also renamed all three species as follows: N. angulatus as Archaias spirans (p. 190), N. aduncus as Helenis spatosus (p. 194), and N. orbiculus as Ilotes rotalitus (p. 198). All three of De Montfort’s species above are considered to be synonymous with Archaias angulatus (Rögl and Hansen 1984).